



������4)�ٽ�����ϸ����ʿ�ԭ��ɱ������ϸ���Ĺ��ܣ����������MHC���ԭ��FcR�����йء�IL-4��GM-CSF��IL-3��LPS��Эͬ���á�IL-4���յ�����Ѫ����ϸ������G-CSF��M-CSF����ǿ������ϸ���鵼�����ɡ�ɱ�˻��Ժ�ADCC���á�IL-4��С�����ϸ���������ӣ����ٽ�IL-1ra�����������Ƶ���ϸ��IL-1��TNF��IL-6�IJ�����
������5)ЭͬCSF�̼���Ѫϸ������ֳ����G-CSFЭͬ��ǿ��ϸ�������γɣ�Эͬ��ϸ�������أ�EPO)��ǿBFU-E���γɡ�
����IL-4��Ϊ�������ߵ��ڼ��ѽ�������ٴ����顣���⣬����ʼ������������ȱ��֢���ٴ����顣�������ڡ������֤ʵIL-4��������IL-1��IL-6��TNF���ڣ����ٽ�IL-1ra ���������Ӧ��IL-4����Ϊ���ư�Ѫ֢�ݿ��ṩһ���µķ�����
����(��)IL-5
����1980��Takatsu�ȷ�����Tϸ����������Һ�У�����һ�����������Tϸ��������Эͬ����������ԭ�Ŀ���Ӧ�𣬳�ΪTϸ��������ӣ�t cell replacing factor,TRF)�������������Ӷ�Bϸ���͆�������ϸ����ֳ���ֻ�����Ҫ�������ã�����Bϸ������������-��b cell growth factor-��,BCGF-��),IgA��ǿ���ӣ�IgA-enhancing factor,IgA-EF)����������ϸ������̼����ӣ�eosinophil colony-stimulating factor,Eo-CSF)����������ϸ���ֻ����ӣ�eosinophil differentiation factor,EDF)��1986��ͳһ����Ϊ��ϸ������-5��interleukin 5,IL-5)��
����1.IL-5�IJ�����������IL-5��Ҫ�ɻTϸ����������С������Th2��Ⱥϸ��������
����2.IL-5�ķ��ӽṹ�ͻ���1986��Kinashi���IL-5cDNA��¡��С��IL-5��133������л���ɣ���21��������ź��ģ�����IL-5���Ӻ���112������л������ķ�����12��15kDa����3���ǻ���λ�㣬�ǻ����������Ϊ18kDa���ǻ�������IL-5���Ա����Լ�����Ӧ����Ľ������Ҫ���á�С��IL-5ͨ���Զ�������ӵĶ�������ʽ���ڣ�������Ϊ45kDa���˵�IL-5��134������л���ɣ���22������л��ź��ģ�2���ǻ����㣬�˺�С��IL-5����ֱ�λ�ڵ�5�ź͵�11��Ⱦɫ�壬��IL-3��IL-4��GM-CSF����Ѫ���ӵĻ��������������˺���IL-5�ڰ�����ˮƽ����70%��ͬԴ�ԣ�����ѧ�����н��淴Ӧ��
����3.IL-5���塡С��IL-5�ɦ��ͦ���������ɡ�������p60����415�������ᣬ�ǵ��ף��ȵ�����17�������ᣬ��Ĥ����322������л�����Ĥ��22������������54��������л��������������IL-5Ϊ�������������źŵ�ת����������p130�����������IL-5���������ͬ��ɸ��������塣С��IL-5R����IL-3R�Ħ�����AIC2B�������)��ͬ����IL-5R��������IL-3R������GM-CSFR���ǹ�ͬ�ġ�����mRNA���ӵIJ�ͬ���ѷ����ж��ֿ�����С��IL-5R����������һ�ֿ�����IL-5��Ĥ���IL-5R�Ľ�ϡ��˺���IL-5R������79%��ͬԴ�ԡ�
����4.IL-5������ѧ���ԡ�������IL��ȣ�IL-5����ѧ������������Խ�խ��
������1)С��IL-5�ٽ���ԭ�̼���Bϸ���ֻ�Ϊ����ϳ�ϸ������Ҫ�����ڽ���ϸ����ֳ���ڵ�Bϸ���������ӻBϸ��IL-2R�ı��IL-5�����ִ̼���������IL-6�������ƣ���IL-5ֻ������Bϸ���̼����խ��ʱ���ڡ�
������2)�ٽ�IgA�ϳɣ�����������ǣ�����ΪIgA�������������ӣ�ʹmIgM ����Bϸ���ֻ�ΪmIgA����Bϸ������������IgA��Bϸ�����ٽ�����ֳ�ͷֻ�����Ϊ����IgA�Ľ�ϸ����IL-4��ЭͬIL-5�ٽ�IgA�ϳɵ����á�IL-5��IgM�ķ���Ҳ�дٽ����á�ҽѧȫ����www.med126.com
������3)ЭͬConA��IL-2�յ�������ɱ����Tϸ��ǰ�壨CTPp)�ֻ�ΪCTL��
������4)��������������ϸ�����ӳ�������������ϸ���Ĵ��ʱ�䣬�̼��˺�С����������ϸ���Ĺ��ܣ��յ���������ϸ���ķֻ���
��������)IL-6
����1980�귢�ֳ���άϸ����Poly I-C�̼����ܲ���һ�����Ʋ������Ƶ�ϸ�����ӣ���Ϊ��2�����أ�IFN-��2)���Ժ���о����δ��֤ʵ�������ӵ�ֱ�ӿ��������ã�������������������ѧ���ܣ�����ʵ��ϵͳ���ܵIJ�ͬ����������Ϊ�ӽ���/��ϸ�����������ӣ�hybri-doma/plasmacytoma growth factor,HPGF),Bϸ���ֻ����ӣ�B cell differentiation factor,BCDF),Bϸ���̼�����-2��b cell stimulatory factor 2,BSF-2),26kDa,��ϸ����Tϸ���ֻ����ӣ�cytolytic T cell differentiation factor,CDF)��ϸ���̼����ӣ�hepatocyte stimu-lating factor,HSF)�ȡ�1986��ͳһ������ϸ������6��interleukin 6,IL-6)��
����1.IL-6�IJ������ܰ�����ijЩ���ܰ���ϸ�����ɲ���IL-6����
������1)Tϸ����Tϸ������IL-6�����ھ���ϸ����PMA����ԭ���ϸ���̼���Ӧ��Tϸ����¡���Լ�HTLV-I��Ⱦ��Tϸ��ϵ�Ⱦ��ɷ���IL-6��
������2)Bϸ������SAC�̼������Bϸ����
������3)����ϸ����LPS�̼�����ϸ������IL-6��ijЩ����ϸ��ϵ��P388D1Ҳ�ɷ���IL-6��
������4)����άϸ�������Է�����IL-6���������ӻ�̼�����IL-1��TNF��PDGF��IFN-�¡�PolyI-C��A23187��PMA�ȿɴٽ�IL-6�IJ�����
������5)��С��ϵĤϸ��������ϸ������Ƥϸ������һ�����������¾��ɲ���IL-6�����⣬����ϸ����ϸ��ϵ��MG63����������T24���װ���A549�ΰ���7860������SK-MG-4����ĸϸ������U373��״ϸ����������ճҺ��ϸ��������ϸ����Ҳ�ܷ���IL-6��������ִ���ǰҶ�е����ݡ���״ϸ����folliculostellate)�ɲ���IL-6���������Ѫ֢ʱLPS�̼�����GH��ACTH�ȼ���ˮƽ�����йء�
����IL-1��TNF��IFN-�¡�PDGF��LPS��Poly ��-C��A23187��PMA�ȶ�IL-6�IJ����������������á�
����2.IL-6�ķ��ӽṹ�ͻ���1985��Kishimoto�ȴ���Tϸ�������Ȼ��IL-6cDNA��¡�ɹ�����IL-6������С����65%ͬԴ�ԡ���IL-6����λ�ڵ�7��Ⱦɫ�壬��Լ5kb,��5�������Ӻ�4���ں��ӡ�
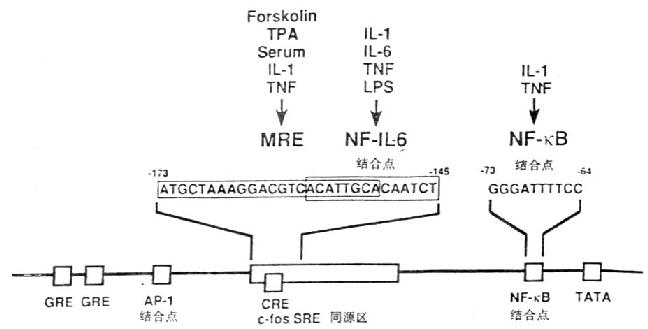
ͼ4-1��IL-6����Ĺ��ܵ�����
������IL-6�����ܵ����������д����Ŷ���ת¼����Ԫ����transcriptional control element)������Ƥ�ʼ��ط�ӦԪ����glucocorticoid responsive elements,GRE)��AP-1���λ�㡢c-fosѪ�巴ӦԪ��ͬԴ�c-fos serum responsive element homology,c-fos SRE homology)��cAMP��ӦԪ����cycli AMp responsive element,CRE)��NF-��B���λ�㡣IL-1��TNF��ϸ�����ӿ�ʹIL-6�����Ӻܿ췢��һ���ԵĻ��IL-1��Ӧ��Ԫ����IL-6��������-180/-123��IL-6�����ӣ�NF-IL-6)ʶ��һ�������14bp,ACATTGCACAATCT���෴ӦԪ����multi-response element,MRE)λ��c-fos SREͬԴ���ڣ���������IL-1��TNF��forskolin��PMA�յ�IL-6�����йأ���IL-1��TNF�̼�IL-6�����йص�NF-��Bλ��TATA�е����Ρ�
������IL-6������212��������л���ɣ�����28��������л����ź����У�����IL-6Ϊ184������л���������26kDa��IL-6������4����������C�ˣ�175��181λ������)�����ϵ�����ɣ�����179λ�������л�����������Ľ�Ϸdz���Ҫ���������ǻ�������ѧ���Թ��ܲ��DZ��裬N��23��������л��䲻ֱ����IL-6����ѧ�����йأ���������IL-6����������ȶ����á���IL-6������������С��IL-6��42%ͬԴ�ԣ��˵�IL-6��С��ijЩϸ���д̼����á�IL-6��G-CSF��IFN-���нϸ�ͬԴ�ԣ��Թ�����Ѫϸ����������Ѫ��ϸ����ijЩ����Ҳ������֮����
����3.IL-6�����塡Ŀǰ��֪��IL-6R�����ɳ�֮ΪIL-6������嵰�ף�IL-6binding receptor protein)�ͳ�Ϊ�ź�ת�����ף�signal-transducing protein)��gp130����ɣ�ϰ����ǰ�߳�֮IL-6R��
������1)IL-6R��CD126)����IL-6R��468����������ɣ��г�N��19��������л���ij��������449�����ᣬ��Ĥ��������Ĥ���Ͱ������ֱ�Ϊ339��28��82�������ᣬ������Ϊ80kDa��6��N�ǻ���λ�㡣��Ĥ����һ��Ig������C2��Լ100������)��2��������ά��ϵ��ṹ������100������)��1��ϸ�����������ͬԴ������ɣ����ߺ�4�����ص�Cys��һ��WSXWS�ṹ������IL-6R��IL-6���Ϊ��������IL-6R�ֲ����ܰ���ϸ���ͷ��ܰ���ϸ������Bϸ����EBVת��Bϸ���������ܰ�ĸϸ����Ѫ��ϸ����������ϸ������ֹTϸ������ϸ��������ϸ��������������Ѫ����AML)ϸ�����ȸ�ϸ����ϸ���ȡ�
������2)gp130(CDw130)��������Ϊ130kDa���ǵ��ף�����14��DZ��N-�ǻ���λ�㣬��Ĥ��������Ĥ���Ͱ������ֱ���597��22��277�������ᡣ��Ĥ������1��IgC2����6��������ά��ϵ��Ľṹ�����еڶ����͵������ṹ��֮����4�����ص�Cys��WSXWS�ṹ�������γ�1��ϸ�������������ṹ�����Ľṹ��gp130����ֱ�������IL-6��ϣ�����������£�IL-6��IL-6�����Ϻ�ʹIL-6R�Ĺ������仯��Ѹ��������gp130���ӽ�ϣ��γɸ������Ľ��λ�㣬��ͨ��gp130�ǵ�λ�����źš��˺�С��gp130�ڰ�����ˮƽ����77%��ͬԴ�ԡ�תȾgp130cDNAС��pro-Bϸ����IL-6/sIL-6R������̼��¿ɴ�����ֳ�źš�С������ע��IL-6������gp130mRNA�ı��Ŀǰ��֤ʵ��gp130�����IL-6�����������⣬Ҳ�ǰ�Ѫ���������ӣ�LIF)��������M��OSM)����״��Ӫ�����ӣ�CNTF)��IL-11�����������õ��ǵ�λ��
������3)�ź�ת����gp130��IL-6/IL-6R�������Ϻ̼�gp130���ڲ��ַ����Ұ������ữ��Ŀǰ���ڲ���˹��̵��Ұ��ᵰ��ø�����û���������ᰱ�ἤø�������̶�����˿����/�հ��ἤø��˿��ԭ��ĵ���ø��mitogen activated protein kinase,MAPK)�ļ��ʹNF-IL-6��˿������հ������ữ��������Ӷ��ٽ���Ӧ����Ļ��
������4)sIL-6R������������������������ϸ��ϵU266�������壬PHA���PBMC�Լ�HTLV-I����ϸ��Ҳ�ܷ���sIL-6R��������Ϊ50kDa���÷�ת¼PCR��������ϸ��������ϸ���о����������sIL-6r mRNA,���з���������Ĥ���������Ӧ��������һ�¡�sIL-6Ҳ�ɴ�Ĥ��ϵ�sIL-6R��mIL-6)���������sIL-6R��IL-12p40�ǻ����и߶�ͬԴ�ԣ���IL-6��IL-12��p35�ǻ����и߶�ͬԴ����˿����Ʋ�������IL-6/sIL-6R�������IL-12���ӿ���Ҳͨ��������pg130����������ϸ����������������ϸ���������岻ͬ��sIL-6R���IL-6�����ϸ��Ĥ����gp130��ϣ���ǿIL-6�Ĵ̼����ԡ���������gp130��sgp130)������sIL-6R/IL-6������Ļ��ԡ�sIL-6Rˮƽ��������ijЩ���������Լ����йء�




